

SUMMARY
Invariant natural killer T (iNKT) cell development in the thymus depends on T cell receptor recognition of CD1d ligand on CD4/CD8 double-positive thymocytes VSports注册入口. We previously reported that B7-CD28 co-stimulation is required for thymic iNKT cell development, but the cellular and molecular mechanisms underlying this co-stimulatory requirement are not understood. Here we report that CD28 expression on CD1d-expressing antigen-presenting T cells is required for thymic iNKT cell development. Mechanistically, antigen-presenting T cells provide co-stimulation through an unconventional mechanism, acquiring B7 molecules via CD28-dependent trogocytosis from B7-expressing thymic epithelial cells, dendritic cells, and B cells and providing critical B7 co-stimulation to developing iNKT cells. Thus, the present study demonstrates a mechanism of B7 co-stimulation in thymic T cell development by antigen-presenting T cells.
In brief
Invariant natural killer T (iNKT) cells mediate various immune responses including cancer immunity. Watanabe et al. show that CD28 expression on antigen-presenting T cells (APTCs) is required for iNKT cell development. APTCs acquire B7 from other B7-expressing cells via CD28-dependent trogocytosis and provide critical B7 co-stimulation for iNKT cell development V体育官网入口.
"V体育官网" Graphical Abstract

"V体育安卓版" INTRODUCTION
Invariant natural killer T (iNKT) cells are a lineage of αβ-T cell receptor (TCR) cells that express an invariant TCRα (Vα14-Jα18 in mouse and Vα24-Jα18 in human) and possess both innate and adaptive characteristics VSports在线直播. 1–3 It is well appreciated that iNKT cells play important roles in regulating immune responses, including responses to infection, anti-tumor responses, and autoimmune responses. 1–3 Development of iNKT cells occurs in the thymus and is dependent on TCR recognition of glycolipid/major histocompatibility complex (MHC) class I-like CD1d complexes presented on CD4/CD8 double-positive (DP) thymocytes. 4–7 In contrast to conventional CD4+ helper and CD8+ killer T cell lineages, which develop in the thymus as phenotypically naive T cells, iNKT cells mature in the thymus with an effector/memory phenotype and exhibit effector functions rapidly upon activation, including effector cytokine production and cytolytic activity. 1–3.
The molecular and cellular requirements for iNKT cell development are substantially distinct from those of conventional lineage T cells. Positive selection of iNKT cells requires antigen presentation by DP thymocytes expressing CD1d,4–7 in contrast to positive selection of conventional T cells, which requires recognition of peptide presented in association with MHC class I or class II molecules expressed by cortical thymic epithelial cells (cTECs). 8 Homophilic SLAM receptor interaction and SLAM receptor associated protein (SAP),9–11 transcription factor PLZF,12,13 and c-myc14,15 are critical for iNKT cell but dispensable for conventional T cell development V体育2025版.
Recent studies have demonstrated that mature thymic iNKT cells can be classified into three distinct lineages based on T helper subset-analogous expression of transcription factors and cytokine production profiles: T-bet-expressing IFN-γ-producing NKT1, GATA3-expressing IL-4-producing NKT2, and RORγt-expressing IL-17-producing NKT17. 16,17 Each iNKT subset plays important roles in conventional T cell development through effector cytokine production that directly affects developing T cells or indirectly modulates thymic medullary epithelial cells (mTECs), as well as exerting distinct effects on peripheral immune responses. 3,18,19 VSports.
B7-CD28 co-stimulation is a well characterized co-stimulatory pathway for many aspects of peripheral T cell immune responses.20,21 In thymic T cell development, the role of B7-CD28 co-stimulation for Treg cell development is well established.22–24 B7-CD28 co-stimulation has also been shown to play a role in deletion of thymic CD8αα IEL precursors.25 We recently carried out an analysis of B7-CD28 requirements for distinct mediators of central tolerance during thymic development26 and found that B7-CD28 co-stimulation is involved in both clonal deletion of conventional T cells and in generation of Treg cells specific for self-tissue-restricted antigens.26 We further demonstrated that these two functions of B7-CD28 co-stimulation have distinct requirements for both cell type-specific B7 expression and cytoplasmic domain-mediated CD28 signaling.26
We and others previously reported an important role of B7-CD28 co-stimulation in thymic iNKT cell development.27–29 However, those studies did not address either the role of B7-CD28 co-stimulation in iNKT subset development or the underlying cellular and molecular mechanisms of B7-CD28 co-stimulation for iNKT cell development. Here we report that B7-CD28 co-stimulation is critical for development of all iNKT subsets and, unexpectedly, that the cellular and molecular mechanisms underlying B7-CD28 co-stimulation for thymic iNKT cell development are distinct from the previously characterized mode of B7-CD28 co-stimulation. CD28 expression by CD1d-positive antigen-presenting T cells is required for acquisition of cell surface B7 by trogocytosis from B7-expressing thymic epithelial cells (TECs), dendritic cells (DCs), and B cells and for providing critical B7 co-stimulation to developing iNKT cells. Our findings thus identify and characterize an unconventional mechanism mediating co-stimulation in thymic T cell development.
RESULTS
V体育ios版 - B7-CD28 co-stimulation is important for thymic development of all iNKT subsets
We first examined the role of B7-CD28 co-stimulation in iNKT subset development in multiple mouse strains. Thymic iNKT cells were detected by PBS57:CD1d-tetramer binding followed by sequential gating strategy (Figure S1A), and NKT subsets were defined by expression of PLZF, T-bet, and RORγt (Figure 1A). NKT1 is dominant in B6 thymus, and NKT2 is dominant in BALB, while CB6F1 showed an intermediate phenotype (Figures 1B and S1). For all genetic backgrounds, absence of B7 (Figure 1) or CD28 (Figure S1B) resulted in significant decrease of all iNKT subsets (Figure 1B). Cytokine-producing capability of the residual iNKT subsets that developed in B7-CD28-deficient mice was assessed after 4-h stimulation with PMA + ionomycin. The proportion of cytokine-producing cells within each iNKT subset was overall normal or slightly reduced relative to wild-type controls, with statistically significant decreases in frequency of IFNγ production by NKT1 and of IL-17 production by NKT17 (Figure 1C upper panel), while the number of cytokine-producing iNKT cells in all subsets, as a proportion of total thymocytes, was significantly and substantially decreased (Figure 1C lower panel).
Figure 1. B7-CD28 co-stimulation is critical for all iNKT cell subset development.
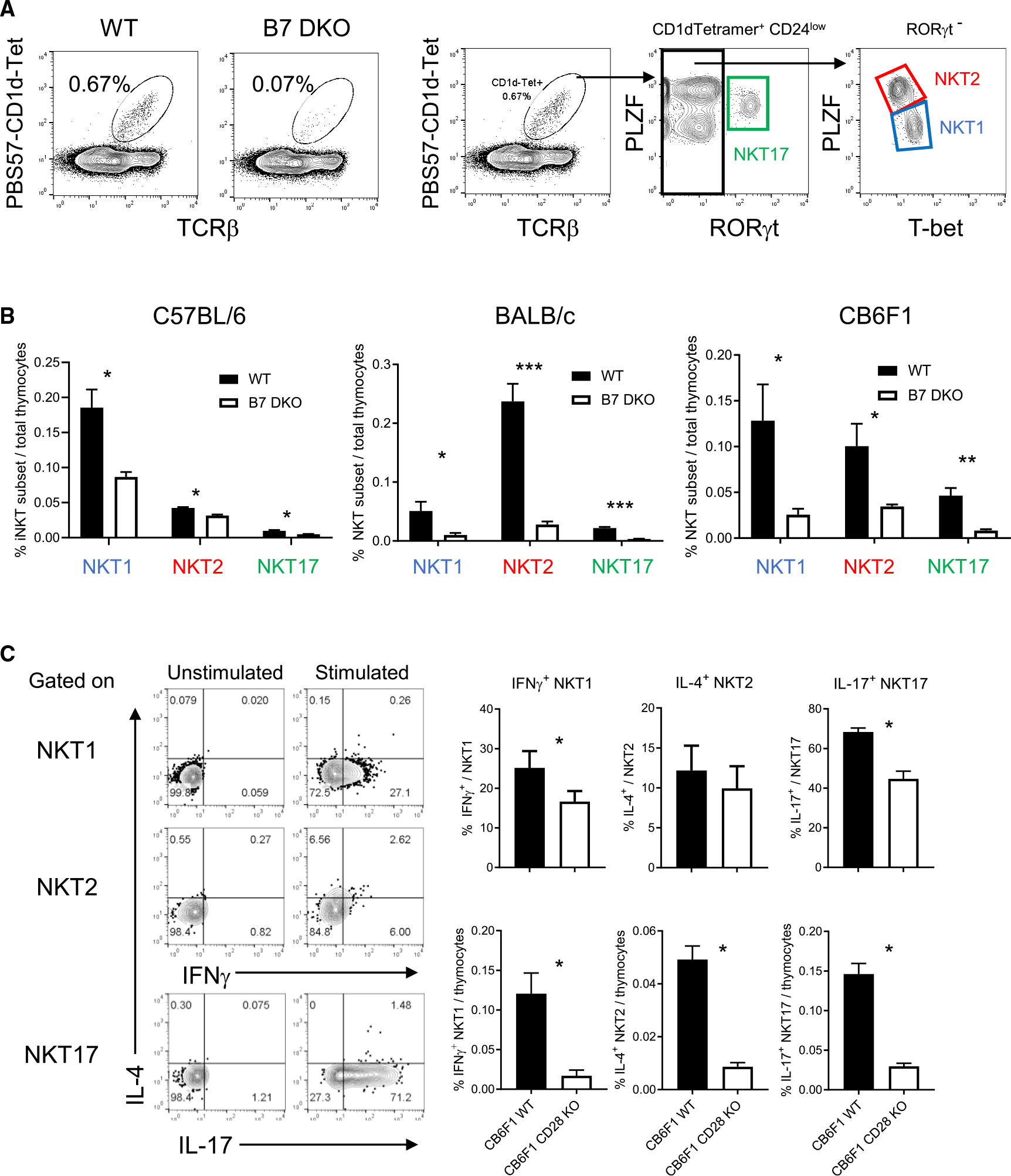
(A) Thymic iNKT cells were defined as B220−, PBS57-CD1d-tetramer binding TCRβ+ cell population. Gating strategy to define NKT1, NKT2, and NKT17 subsets within CD24low iNKT cells.
(B) iNKT cell subset in C57BL/6 (B6), BALB/c, and CB6F1 strains. B6 WT = 6, B6 B7 DKO = 6, BALB/c WT n = 7, BALB/c B7 DKO n = 7, CB6F1 WT n = 4, CB6F1 B7 DKO n = 4. Data are pooled results of three independent experiments.
(C) Cytokine production of each iNKT cell subset. Total CB6F1 thymocytes were stimulated with PMA and ionomycin, and intracellular cytokine staining was performed. Left panel: representative intracellular cytokine staining pattern of each iNKT cell subset defined as in (A). Upper right panel: frequency of indicated cytokine staining positive population per each iNKT cell subset. Lower right panel: frequency of indicated cytokine positive iNKT cell subset population per total thymocytes. CB6F1 WT n = 4, CB6F1 CD28 KO n = 4. Data are pooled results of three independent experiments.
(B and C) Data are mean ± SEM. Statistical difference between groups were calculated with Student’s t test. *p < 0.05, **p < 0.01, ***p < 0.001. See also Figure S1.
We next determined whether, in addition to affecting the number of developing iNKT cells, B7-CD28 co-stimulation plays a role in selection of the TCR repertoire of iNKT cells. It is known that mouse thymic iNKT cells express an invariant TCRα in association with preferentially expressed TCR Vβs (Vβ8, Vβ2, Vβ7), and that iNKT subsets show substantially different patterns in usage of these Vβs.17 We assessed TCR Vβ expression by single-cell 5′ RNA-seq in BALB/c wild-type (WT) and B7 double knockout (DKO) iNKT cells. TCR Vβ usage was significantly altered in B7 DKO iNKT cells, and these effects were selective for individual iNKT subsets (Figure S1C). These results demonstrated a critical role of B7-CD28 co-stimulation for development of all iNKT subsets and for TCR repertoire selection in these subsets.
Strong B7-CD28 interaction occurs during early stages of iNKT cell development
To elucidate underlying cellular and molecular mechanisms of B7 co-stimulation, we first sought to assess B7-CD28 interaction over the course of iNKT cell development. To this end, we defined iNKT cell developmental stages using CD44, CD24, and CD69 expression (Figure 2A). Since higher CD44 expression is a hallmark of mature iNKT cells, we first posited that CD44low cells represent developing iNKT cells, while CD44high represents mature thymic iNKT cells.18 Within the CD44low fraction, we analyzed CD24 (HSA), for which decreasing expression level correlates with increasing maturation of thymic iNKT cells, and CD69, which is an indicator of TCR signaling during thymic iNKT positive selection.18 We postulated a developmental sequence starting with (1) CD44lowCD24highCD69−, followed by (2) CD44lowCD24highCD69+, (3) CD44lowCD24intCD69+, (4) CD44lowCD24lowCD69−, and then (5) CD44high. This sequence of developmental stages was confirmed by comparing expression level of Rag2-GFP as a molecular timer for residency time following positive selection and termination of Rag2 gene expression30 (Figure 2B). We defined the stages analyzed in this study as (1) preselection (prior to TCR signaling), (2) stage 1, (3) stage 2, (4) stage 3, and (5) mature, as shown in Figure 2C. To evaluate thymic localization of each stage of iNKT, CCR7 expression was analyzed. CCR7 is induced on positively selected thymocytes, and its expression is important for cortical to medullary migration.31–33 CD69 and CCR7 expression were inversely correlated during early stages of iNKT development. CD69 was upregulated in stage 1 relative to preselection iNKT and remained high in stage 2 but was downregulated in stage 3 (Figure 2B), while CCR7 was not expressed in stage 1, was upregulated in stage 2, and increased in expression at stage 3 (Figure 2D). Egr2, an early TCR signal response gene shown to be important for NKT cell development, showed kinetics of expression similar to that of CD69 (Figure S2A). These results suggested that stage 1 (CD69+CCR7−) and stage 2 (CD69+ CCR7low) iNKT cells were under positive selection by TCR signaling and localized to the cortex, while stage 3 (CD69−CCR7high) iNKT cells might be migrating and/or have migrated to the medulla as a result of higher CCR7 expression31,33 (Figures 2B and 2D).
Figure 2. Developmental iNKT cell stages and analysis of CD28 expression level.

(A) Gating strategy to identify iNKT cell sub-populations based on CD44, CD24, and CD69 expression.
(B) B6 Rag2-GFP expression level of each iNKT cell sub-population. n = 3. Data are representative of two independent experiments.
(C) Defined developmental sequence of iNKT cell sub-populations.
(D) CCR7 expression pattern at each developmental stage. n = 5. Data are pooled result of independent two experiments.
(E) CD28 expression level (MFI) of each developmental stage in B6 WT, B7 DKO, and CD28 KO strains.
(F) Difference of MFI (ΔMFI) between B7 DKO and WT was calculated at each stage.
(E and F) Each group n = 3. Data are representative of three independent experiments. (B, D–F) Data are mean ± SEM. See also Figure S2.
We evaluated the presence of ongoing B7-CD28 interaction at each developmental stage by measuring B7-dependent downregulation of cell surface CD28, as previously reported.34 In WT mice, CD28 was upregulated upon positive selection (stage 1), and its level subsequently decreased during maturation (Figure 2E). To assess B7-dependent CD28 downregulation, CD28 expression level at each stage of iNKT cell development was measured in B7 DKO mice. After CD28 upregulation at stage 1, its expression level was further upregulated in B7 DKO thymocytes through stage 2, after which expression decreased (Figure 2E). At each stage of differentiation, CD28 expression was greater in B7 DKO thymocytes than in WT. As a quantitative index of B7-CD28 interaction, the difference in mean fluorescence intensity (MFI) between B7 DKO and WT was plotted. This evaluation indicated that B7-CD28 interaction, as reflected by B7-dependent downregulation of CD28,34 had already been initiated at stage 1, peaked at stage 2, and decreased at later stages of development (Figure 2F). These results indicated that strong CD28 upregulation and B7-CD28 interaction occurs during positive selection, likely in the thymic cortex.
B7-CD28 co-stimulation is critical for the developmental transition from stage 1 to stage 3 of iNKT cells at an early stage of thymic selection
To determine the effect of B7-CD28 co-stimulation on iNKT cell development, cell numbers at each developmental stage were analyzed in WT and in co-stimulation-deficient thymus. iNKT cell numbers were comparable in WT and co-stimulation-deficient thymus at preselection (CD24highCD44lowCD69−) and stage 1 (CD24highCD44lowCD69+), while cell numbers were significantly reduced in co-stimulation-deficient thymus at stage 2 (CD24intCD44lowCD69+), and markedly reduced in stage 3 (CD24lowCD44lowCD69−) and mature (CD24lowCD44high) iNKT cells (Figure 3A). These results indicated that B7-CD28 co-stimulation is critical for the developmental transition from stage 1 to stage 2 and stage 3 of iNKT cell development.
Figure 3. Effect of absence of B7 co-stimulation on developing iNKT cell sub-populations.
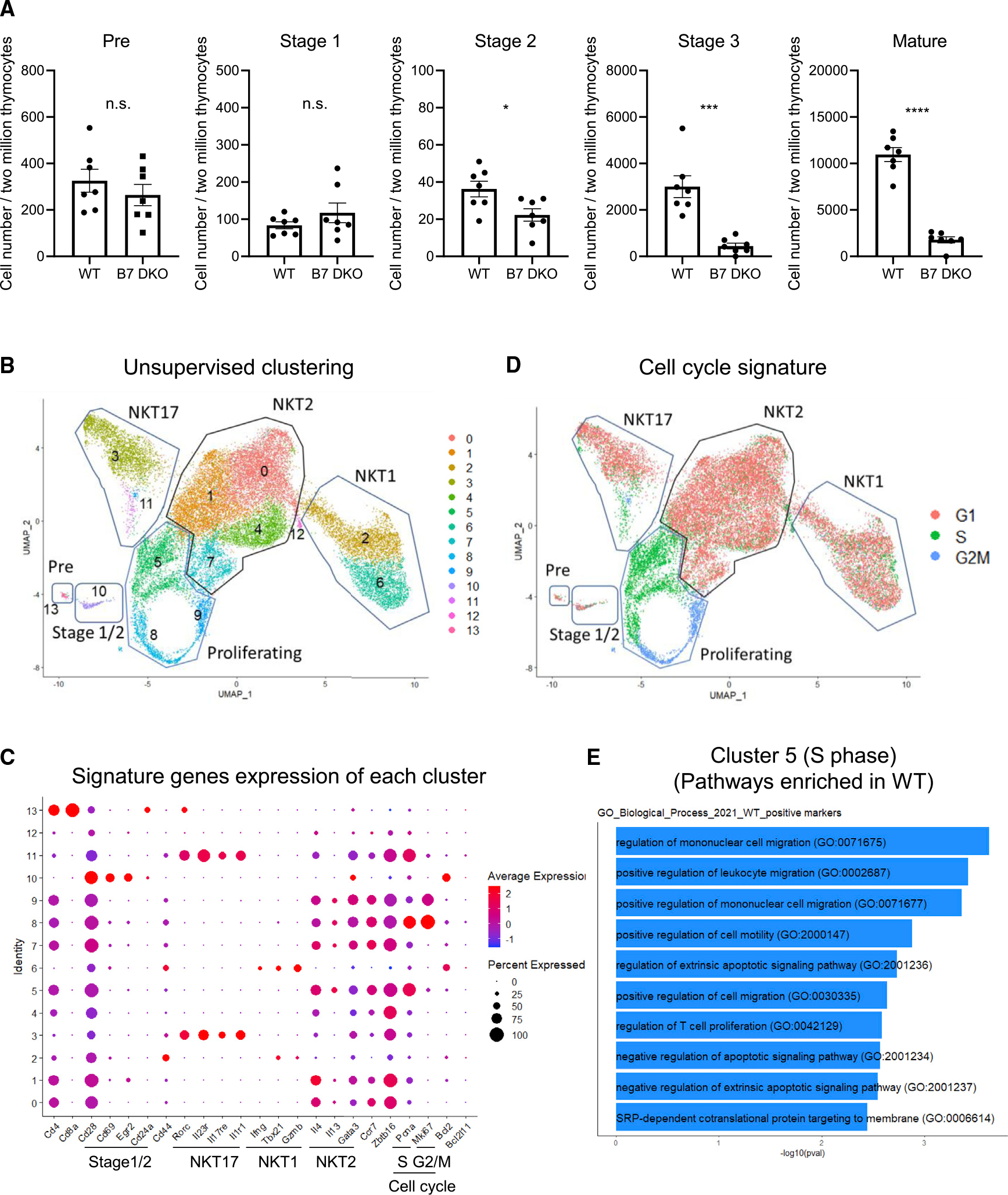
(A) Cell number at each developmental stage. BALB/c thymic iNKT cells were first enriched by magnetic-activated cell sorting with CD1d-tetramer and then analyzed by fluorescence-activated cell sorting (FACS). Data are expressed as cell numbers in each developmental stage within the tetramer-enriched iNKT cell population. Each group n = 7. Data are pooled results of three independent experiments.
(B) Uniform manifold approximation and projection (UMAP) of six independent scRNA-seq data sets from BALB/c thymic iNKT cells (WT n = 3, B7 DKO n = 3) integrated using Seurat.
(C) Scaled expression of selected signature genes for each cluster colored by average expression of each gene in each cluster. Dot size represents the percentage of cells in each cluster.
(D) Cell cycle profiling of each cluster.
(E) Gene ontology (GO) enrichment analysis. Differentially expressed genes between WT and B7 DKO for cluster 5 were identified using Wilcoxon rank-sum test. GO_Biological_Process_2021 database was used for GP enrichment analysis.
(A) Data are mean ± SEM. Statistical differences between groups were calculated with Student’s t test. **p < 0.01, ***p < 0.001. See also Figure S3.
The underlying molecular mechanism(s) mediating CD28 co-stimulation for thymic iNKT cell development are not well understood. For global analysis of developing thymic iNKT cells, we performed transcriptomic characterization by scRNAseq of B7 WT and DKO thymic iNKT cells. Since B7-CD28 deficiency resulted in larger reduction in NKT cell number in BALB/c compared with B6, and BALB/c has more balanced composition of NKT1/2/17 subsets compared with B6 (dominated by NKT1), BALB/c mice were used for scRNAseq analysis. To allow concurrent analysis of sufficient numbers of both the quantitatively predominant CD44high mature iNKT cells and the much less frequent CD44low immature iNKT cells, we flow cytometrically sorted mature (CD44high) and immature (CD44low) iNKT cells separately; then we mixed these two populations in equal proportion for scRNAseq analysis. Unsupervised clustering analysis identified 13 clusters based on differential patterns of gene expression. These corresponded to a preselection cluster, a stage 1/2 signature (CD28highCD69highEgr1/2high) cluster, proliferating clusters, and NKT1, NKT2, and NKT17 clusters (Figures 3B and 3C). B7 WT and B7 DKO showed similar cluster composition (Figure S3A). Clusters 5, 8, and 9 were classified as proliferating, with cluster 5 expressing an S phase cell cycle signature and clusters 8 and 9 expressing G2/M signatures (Figure 3C). The cell cycle signatures of the clusters presented in Figure 3B are shown in Figure 3D. Since a clonal burst of developing iNKT cells occurs after positive selection and before the mature iNKT cell stage,35 and since we observed that a decrease in B7 DKO versus WT cell numbers became apparent in the transition from stage 2 to stage 3, we hypothesized that lack of B7 co-stimulation might impact gene expression related to proliferation and cell cycle in developing iNKT cells. In cluster 5 (S phase cell cycle signature), pathways including T cell proliferation (GO: 0042129) and negative regulation of apoptotic signaling pathway (GO: 2001234) were upregulated in WT relative to B7 DKO (Figure 3E), suggesting a role for B7-CD28 co-stimulation in regulating proliferation and apoptosis in proliferating thymic NKT cells. Of note, in differentially expressed gene analysis, Ckb (creatine kinase B) expression was consistently lower in B7 DKO NKT, while Crip1 (cysteine rich protein 1) was higher in B7 DKO NKT (Figures S3B and S3C), suggesting these as potential candidate genes in future analysis.
CD1d-expressing thymocytes acquire B7 via CD28-dependent trogocytosis (VSports手机版)
We next sought to determine the cell type(s) involved in B7 presentation to iNKT cells during development. Since CD1d-expressing DP thymocytes are uniquely required as antigen-presenting cells during iNKT cell selection,4–7 two possible mechanisms of B7 co-stimulation appeared possible, both unconventional. It was possible that B7 co-stimulation is provided in cis by the same CD1d-expressing DP T cells that function in positive selection of iNKT cells. This would be unconventional in that B7 expression and co-stimulation by DP thymocytes has not been previously identified. Alternatively, it was possible that co-stimulation is provided in trans, i.e., by a cell distinct from the CD1d-expressing DP thymocytes that are required to provide the first or TCR-mediated signal for iNKT cell development. Potential sources of B7 co-stimulation in this case might include TECs, DCs, B cells, and single-positive (SP) T cells, noting that there is no requirement for CD1d expression on any of these cells for iNKT cell development.5,7,35 Either case would represent a quite non-conventional mode of B7-CD28 co-stimulation.
Flow cytometric analysis demonstrated that cell surface B7 is indeed expressed on thymic DP and SP T cells (Figures 4A and S4A), in addition to the expected expression of B7 on TECs, DCs, and B cells (Figure S4B). To address cellular requirements for B7 co-stimulation in iNKT cell development, we utilized a B7 conditional knockout (KO) strategy.36 Thymic iNKT cell development in B7flox mice is comparable to that of WT B6 mice (Figure S4C). B7 was deleted from specific cell types of B7flox mice, including TECs, DCs, B cells, or T cells by using Foxn1-Cre, CD11c-Cre, CD19-Cre26 (Figure S4B), or proximal-Lck-Cre, respectively. However, none of these conditional knockout (cKO) strains showed a defect in thymic iNKT cell development (Figures 4B and 4C). In contrast, B7 deletion from all TECs, DCs, and B cells in triple-Cre mice resulted in abolishment of B7-dependent iNKT cell development to a similar level as that observed in B7 complete KO mice (Figure 4B). This result might indicate that B7 on TECs, DCs, and B cells has a redundant capacity to mediate B7 co-stimulation in trans for iNKT cell development. However, on careful evaluation of the cell type specificity of B7 deletion, we noted that B7 expression on peripheral T cells was eliminated in TEC/DC/BC-B7 triple-Cre cKO mice, while B7 expression on T cells was not eliminated by genomic deletion of B7 in T cell-specific B7cKO mice (Figure 4D). It has been reported that B7 can be acquired on peripheral T cells as a result of interaction of T and antigen-presenting cells (APCs) through CD28-dependent trogocytosis.37,38 We therefore tested the possibility that this mechanism could be involved in the observed B7 expression on T cells, using a mixed bone marrow (BM) chimera strategy. Indeed, B7-gene-deficient peripheral T cells derived from B7 DKO BM acquired B7 molecules in vivo when B7-expressing cells co-existed in these mixed chimeras (Figure 4E). This B7 acquisition was CD28 dependent, as B7 was not acquired by CD28 KO T cells (Figure 4F). Notably, CD28-dependent B7 trogocytosis was not restricted to peripheral T cells, since B7 expression on thymocytes, including DP and SP cells, was also observed and was similarly CD28 dependent (Figure 4A). Of interest, truncation of the CD28 cytoplasmic tail did not affect B7 expression on thymocytes, indicating that the CD28 intracellular domain was not required for B7 acquisition (Figure 4G).
Figure 4. Thymocytes acquire and express B7 on cell surface in a CD28-dependent manner.

(A) B7.1 and B7.2 expression on thymocytes in B6 WT, B7 DKO, and CD28 KO mice. Top panel: representative histogram results of at least three independent experiments. Bottom panel: quantification of pooled results for B7.1 and B7.2 MFI from four independent experiments. WT n = 5, CD28 KO n = 5, B7 DKO n = 3.
(B) iNKT cell population in B6 DC, B, and/or TEC-specific B7 conditional KO mice generated by crosses with CD11c-Cre, CD19-Cre, and/or Foxn1-Cre respectively. B7flox n = 9, B7 DKO n = 4, TEC/DC/BC B7 cKO n = 3, TEC B7 cKO n = 5, DC B7 cKO n = 4, BC B7 cKO = 5. Data are pooled results of four independent experiments.
(C) iNKT cell population in T cell-specific B7 conditional KO mice generated by cross with Lck-Cre. B7flox n = 24, B7 DKO n = 26, TEC/DC/BC B7 cKO n = 3, TC B7 cKO n = 5. Data are pooled result of at least three independent experiments.
(D) Cell surface B7.1 expression on peripheral splenic CD4+ T cells in B6 B7 conditional KO strains. Red line indicates B7.1 staining of each cKO mouse. Black line indicates B7.1 staining of B7 DKO mice as negative staining control. Data are representative results of at least three independent experiments.
(E) B7 expression on B6 B7 DKO T cells in mixed bone marrow chimeras with co-existence of B7 WT T cells. Red line indicates B7.1 staining of each cell type. Black line indicates B7.1 staining on each cell type from B7 DKO mice as negative staining control.
(F) B7 expression on peripheral CD4+ T cells in B6 CD28 KO mice. B7.1 expression on splenic CD4+ T cells and CD11c+ dendritic cells is analyzed. Red line indicates B7.1 staining of each cell type. Black line indicates B7.1 staining of each cell type from B7 DKO mice as negative staining control. Data are representative results of at least three independent experiments.
(G) B7 expression on DP thymocytes in B6 CD28-cytoplasmic-region-truncated mutant mice. Data are representative results of two independent experiments. Each group total n = 4.
(A–C) Data are mean ± SEM. Statistical differences between groups were performed with one-way ANOVA followed by multiple comparison. *p < 0.05, **p < 0.01, ****p < 0.0001. See also Figure S4.
"VSports手机版" CD28 expression on CD1d-expressing antigen-presenting T cells is required for NKT cell development
Based on these observations, we hypothesized that CD28-dependent B7 acquisition on T cells is a critical mechanism for providing B7 co-stimulation to developing thymic iNKT cells. This possibility was tested in mixed BM chimeras. One donor bone marrow (BM1) was CD1d KO (CD45.1), and cells derived from this BM were used as a readout for iNKT cell development, since these cells alone would not be able to support iNKT cell development due to lack of CD1d. The second donor bone marrow (BM2) in mixed chimeras was CD1d WT (CD45.2), to test the APC capability of this CD1d-expressing population for supporting iNKT cell development of BM1-derived cells. We tested the requirement for CD28 expression on BM2 antigen-presenting T cells by comparing CD1d WT/CD28 WT (group 2) and CD1d WT/CD28 KO (group 3) bone marrow as BM2. T cells from CD1d WT/CD28 KO bone marrow (group 3, BM2) express CD1d but are not able to acquire B7 due to lack of CD28, while DC and B cells derived from the same BM express both CD1d and B7 (Figures 5A–5C). When BM2 was CD1d KO, iNKT cells did not develop in the BM1 compartment (group 1), while wild-type (CD1d WT/CD28 WT) BM2 cells did support iNKT cell development in the BM1 compartment (group 2). When CD1d WT/CD28 KO BM was tested for APC capability to support iNKT cell development (group 3), the efficiency of iNKT cell development was markedly decreased, to a level similar to that seen in the complete absence of CD28 (group 4) (Figure 5A). We confirmed CD28 KO DP cells express CD1d and SLAM family molecules at a comparable level to those of CD28 WT DP (Figure 5D). It should be noted that in the group 3 BM chimera mice, T cells from BM1 were CD1d KO but CD28 WT, and thus able to acquire B7 through trogocytosis, and that BM2-derived cells other than T cells, including DCs and B cells, expressed both CD1d and B7 (Figure 5C); nevertheless, those cell populations were not sufficient to support B7-CD28 co-stimulation-dependent iNKT cell development (Figure 5A). Consistent with reported results,5 CD1d-expressing SP cells were not required for iNKT cell development, since TCRα KO cells, in which T cell development was arrested at DP stage, were still capable of providing both TCR signal and co-stimulation (Figure S5A). The requirement for CD28 expression on antigen-presenting T cells for iNKT cell development was similarly observed in BALB/c BM chimera mice (Figure S5B). Of interest, the CD28 cytoplasmic tail was dispensable for both B7 acquisition (Figure 4G) and for the ability of CD1d+ T cells to function as APCs in support of B7-dependent iNKT cell development (Figure S5C). Taken together, these results indicated that co-expression of CD1d and CD28 on antigen-presenting DP thymocytes is required for B7 trogocytosis and for providing B7 co-stimulation for iNKT cell differentiation.
Figure 5. CD28 on antigen-presenting T cells is required for B7 co-stimulation in iNKT cell development.

(A) CD28 expression on CD1d-expressing antigen-presenting T cells is required to co-stimulate iNKT cell development. To test the requirement for CD28 expression on antigen-presenting-cells for iNKT cell development, mixed bone marrow (BM) chimera mice on B6 background were generated. In groups 1, 2, and 3, BM1 was CD1dKO (CD45.1) and served as the indicator of iNKT cell development. BM2 (CD45.2) was CD1dKO/CD28WT (group 1), CD1dWT/CD28WT (group 2), or CD1d WT/CD28KO (group 3) as the source of CD1d-expressing APCs. In group 4, irradiated CD28 KO mice were reconstituted with CD28 KO BM to generate chimeras that were completely lacking CD28. 10 weeks after BM reconstitution, iNKT cells in BM1 compartment were analyzed. Group 1, n = 8; group 2, n = 6; group 3, n = 10; group 4 n = 7. Data are pooled results of at least three independent experiments. Data are mean ± SEM. Statistical comparisons between groups were performed with one-way ANOVA followed by multiple comparison. ***p < 0.001,
(B) Solid red line represents B7.1 expression on CD1d+ DP thymocyte (BM2) in BM chimera mice in (A). Black dotted line is anti-B7.1 staining of B7 DKO DP as negative staining control. Data are representative result of at least three independent experiments.
(C) B7 expression pattern on CD1d+ antigen-presenting cells and CD1d− non-antigen-presenting-cells. See also Figure S5.
DISCUSSION
We identify a requirement for B7-CD28 co-stimulation for all iNKT cell subsets, and we describe here the mechanism mediating B7-CD28 co-stimulation and its function in iNKT cell development. We report that thymic iNKT cell development requires B7-CD28 co-stimulation during positive selection and, strikingly, that this co-stimulation is mediated through an unanticipated mechanism. The CD1d-positive antigen-presenting T cells for iNKT cell selection must express CD28 in order for these cells to acquire B7 from B cells, DCs, and TECs by trogocytosis and to provide the functional B7 co-stimulation required for iNKT cell development.
The CD28 co-stimulatory signal pathways responsible for iNKT cell development have not previously been elucidated. iNKT cell development critically depends on transcription factors Egr239 and PLZF.12,13 Nevertheless, Egr2 and PLZF protein expression were not affected in residual iNKT cells in the B7-CD28 co-stimulation-deficient mice (Figure S2A). To understand the role of B7-CD28 co-stimulation in thymic iNKT cell development, we performed single-cell RNA sequencing (scRNA-seq) analysis on developing WT and B7 DKO iNKT cells. In cells expressing an S phase cell cycle signature (Figure 3C, cluster 5), pathways such as T cell proliferation (GO: 0042129) and negative regulation of apoptotic signaling pathway (GO: 2001234) were upregulated in WT relative to B7 DKO (Figure 3E), suggesting a role for B7-CD28 co-stimulation in regulation of proliferation and apoptosis in proliferating thymic iNKT cells. Of interest, Ckb and Crip1 expression was consistently lower or higher in B7 DKO NKT, respectively (Figures S3B and 3C), suggesting these as potential candidate genes in future analysis.
While the CD28 cytoplasmic tail is required for iNKT cell development,28 we found here that the Y170 residue and/or C-terminal proline motif of CD28 cytoplasmic tail were dispensable (Figure S2B), indicating that as yet undefined CD28 cytoplasmic domain(s) are responsible for the CD28 co-stimulation that is required for iNKT cell development. This requirement for CD28 cytoplasmic motifs is similar to that for thymic clonal deletion but different from that for Treg cell generation,26 further defining the distinct CD28 cytoplasmic domain requirements, and presumably downstream signaling pathways, for different pathways of agonist selection. Of note, neither CD28-dependent B7 trogocytosis nor APC capability of DP cells requires the CD28 cytoplasmic tail (Figures 4G and S5C), distinguishing these from other CD28 functions in iNKT and T cell development.28
Positive selection of iNKT cells is mediated by antigen-CD1d complexes on DP thymocytes, while CD1d expression by other cell types is notrequired.5,7,35 This fact led us to seek identification of the cellular requirement for B7 expression in iNKT cell development, since either B7 co-stimulation by antigen-presenting T cells or B7 co-stimulation in trans by non-APCs would differ from previously described modes of B7-CD28 co-stimulation. To this end, we first employed a B7 conditional deletion strategy and found that B7 deletion from either TECs, DCs, B cells, or T cells did not affect iNKT cell development, while combined B7 deletion from TECs, DCs, and B cells abrogated B7-dependent iNKT cell development (Figures 3B and 3C). These observations suggested that B7 co-stimulation might occur in trans by cells distinct from the CD1d-antigen-presenting T cells. However, evaluation of surface B7 expression in each B7 conditional KO strain revealed that in triple-cre mice, with deletion of B7 from TECs, DCs, and B cells, B7 expression on T cells was eliminated, while B7 was expressed at undiminished levels in T cell-specific B7-cKO with genomic deletion of B7. These results suggested the possibility that cell surface B7 might be transferred to T cells from B7-expressing non-T cells and that the acquired B7 on T cells might provide co-stimulation for developing iNKT cells.
It has been reported that peripheral T cells can acquire cell surface molecules that are expressed on APCs through a process termed trogocytosis.40 In thymus of BM chimeras, it was reported that donor BM-derived thymocytes express host-derived MHC,41 indicating acquisition during thymic T cell development. Trogocytosis has been implicated in a number of biological processes, including immune responses, tumor growth, neural development, and pathogen invasion of hosts.40 Through T cell-APC interaction, peripheral T cells can acquire cell surface molecules including MHC, B7, and ICAM-1.37 This B7 acquisition by peripheral T cells is CD28 dependent.37 Indeed, we found that B7 expression on thymocytes was also CD28 dependent, since B7 expression on CD28-deficient thymocytes was completely abrogated (Figure 4A). Interestingly, the CD28 intracellular domain was not required for B7 acquisition on thymocytes (Figure 4G), indicating that CD28 cytoplasmic tail-mediated signals are dispensable for B7 trogocytosis.
Using mixed BM chimeras, we directly tested whether CD28-dependent B7 trogocyotosis is important for iNKT cell development. When CD1d-expressing antigen-presenting T cells in mixed BM chimeras were CD28 deficient, B7 expression on CD28-deficient DP was much reduced, and iNKT cell development was severely impaired. Of note, in these chimeras, cell types that do not function as CD1d-presenting APCs for iNKT cell development, such as TECs, DCs, and B cells, still express wild-type levels of B7, but this B7 expression was not sufficient to support iNKT cell development. These results demonstrated that CD28 expression on CD1d-antigen-presenting T cells is critical both for acquisition of B7 by trogocytosis and to provide B7 co-stimulation for developing iNKT cells.
Models of B7-CD28 co-stimulation have generally concluded that co-stimulation is provided in cis, with the same APCs involved in both Ag/MHC-TCR and B7-CD28 interaction.42 Our results for iNKT cell development also identify a requirement for cis co-stimulation. However, in previously described models, both MHC and co-stimulatory molecules such as B7 are genomically encoded by the same APC. A few reports have suggested that B7 co-stimulation can work in trans, with Ag/MHC and B7 presented by different cells.43 However, those data were based on in vitro experiments under conditions of non-physiological stimulation, and it is not clear whether B7-CD28 co-stimulation can function in trans under in vivo physiological conditions of T cell development and/or peripheral T cell activation.
Our results indicate that B7-CD28 interaction occurs early during iNKT cell positive selection, in cells that express high levels of CD69 and Egr2 and relatively low levels of CCR7 and that may still be in the thymic cortex. While it is thought that cTECs do not express B7, BM-derived cells including DCs, B cells, and macrophages44 are potential cell types involved as sources of B7 for CD28-dependent trogocytosis by DP thymocytes in the thymic cortex. Although B7 expression levels detected on thymocytes by flow cytometry were lower than those on other thymic cells, such as mTECs, DCs, and B cells (Figures 4A and S4B), this lower expression level might in fact be advantageous for iNKT cell development, since we previously reported that high levels of B7 expression negatively impacted thymic iNKT cell development, especially when transgene-encoded B7 was highly expressed in T cells.28
Taken together, our findings elucidated a mechanism of B7-CD28 co-stimulation in T cell development mediated by CD28-dependent B7 trogocytosis by antigen-presenting T cells. B7 expressed by TECs, B cells, and DCs is acquired by CD28 expressed on CD1d-expressing DP thymocytes, which then provide both Ag/MHC for TCR recognition and acquired B7 for CD28-dependent co-stimulation.
Limitations of the study
This study identified a requirement for CD28 expression on DP thymocytes for acquisition and presentation of B7 ligand and for providing B7-CD28 co-stimulation to developing thymic iNKT cells in mouse. Since we utilized CD28 KO mice to demonstrate the importance of CD28 expression on DP cells to acquire B7, we cannot completely exclude the possibility that CD28 KO DP cells as APCs have cell intrinsic defect(s) other than B7 acquisition to support iNKT cell development, despite our inability to identify any such differences. We showed that thymic DCs, B cells, and TECs have redundant roles as sources of B7 for trogocytosis by utilizing B7.1 cKO (B7.2 KO) analysis. This B7.1 cKO system can test the cell type-specific role of B7.1 but cannot address the cell type-specific role of B7.2, if any. It is known that iNKT cell subset development and composition vary among mouse strains. In this regard, we incorporated three mouse strains, C57BL/6, BALB/c, and CB6F1, in this study, but in some analyses, we were unable to analyze all strains due to technical and/or resource limitation.
STAR★METHODS
RESOURCE AVAILABILITY
Lead contact
Further information and request for resources and reagents should be directed to and will be fulfilled by the lead contact, Richard J. Hodes (hodesr@qiuluzeuv.cn).
Materials availability
This study did not generate new unique reagents.
Data and code availability
Single-cell RNA-seq data have been deposited at the NCBI Gene Expression Omnibus (GEO: GSE198970) and are publicly available as of the date of publication. Accession numbers are listed in the key resources table. Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
This paper does not report original code.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
KEY RESOURCES TABLE.
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
|
| ||
| Antibodies | ||
|
| ||
| Anti-mouse CD4-BV510 (clone RM4-5) | BD Bioscience | Cat# 563106; RRID:AB_2687550 |
| Anti-mouse CD44-FITC (clone IM7) | BD Bioscience | Cat# 561859; RRID:AB_10894581 |
| Anti-mouse TCRβ-PE/Cy7 (clone H57-597) | BD Bioscience | Cat# 560729; RRID:AB_1937310 |
| Anti-mouse RORgt-BV786 (clone Q31-378) | BD Bioscience | Cat# 564723; RRID:AB_2738916 |
| Anti-mouse Tbet-BV421 (clone O4-46) | BD Bioscience | Cat# 563318; RRID:AB_2687543 |
| Anti-mouse PLZF-AF647 (clone R17-809) | BD Bioscience | Cat# 563490; RRID:AB_2738238 |
| Anti-mouse CCR7-BV786 (clone 4B12) | BD Bioscience | Cat# 564355; RRID:AB_2738765 |
| Anti-mouse IL-17A-AF488 (clone TC11-18H10) | BD Bioscience | Cat# 560220; RRID:AB_1645194 |
| Anti-mouse IFNγ-FITC (clone XMG1.2) | BD Bioscience | Cat# 554411; RRID:AB_395375 |
| Anti-mouse IL-4-BV421 (clone 11B11) | BD Bioscience | Cat# 562915; RRID:AB_2737889 |
| Anti-mouse NK1.1-APC/eF780 (clone PK136) | Thermo Fisher | Cat# 47-5941-82; RRID:AB_2735070 |
| Anti-mouse CD8α-APC/eF780 (clone 53-6-7) | Thermo Fisher | Cat# 47-0081-82; RRID:AB_469335 |
| Anti-mouse B7.1-APC (clone 16–10A1) | Thermo Fisher | Cat# 17-0801-82; RRID:AB_469417 |
| Anti-mouse B7.2-APC (clone GL1) | Thermo Fisher | Cat# 17-0862-82; RRID:AB_469419 |
| Anti-mouse CD28-APC (clone 37.51) | Thermo Fisher | Cat# 17-0281-82; RRID:AB_469374 |
| Anti-mouse Egr2-APC (clone erongr2) | Thermo Fisher | Cat# 17-6691-82; RRID:AB_11151502 |
| Anti-mouse B220-AF700 (clone RA3-6B2) | BioLegend | Cat# 103232; RRID:AB_493717 |
| Anti-mouse CD24-AF594 (clone M1/69) | BioLegend | Cat# 101834; RRID:AB_2565427 |
| Anti-mouse CD69-BV421 (clone H1.2F3) | BioLegend | Cat# 104545; RRID:AB_2686969 |
|
| ||
| Chemicals, peptides, and recombinant proteins | ||
|
| ||
| Mouse CD1d PBS-57 (PE-Labeled Tetramer) | NIH Tetramer core facility | Cat# 58281 |
| PMA | Sigma | Cat# P8139-1MG |
| Ionomycin | Sigma | Cat# 10634-1MG |
| EasySep PE-positive selection kit II | STEMCELL | Cat# 17684 |
| Foxp3/Transcription factor staining buffer set | Thermo Fisher | Cat# 00-5523-00 |
| GolgiPlug | BD Bioscience | Cat# 555029 |
| GolgiStop | BD Bioscience | Cat# 554724 |
|
| ||
| Deposited data | ||
|
| ||
| Single-cell RNA-seq data | This paper | GEO: GSE198970 |
|
| ||
| Experimental models: Organisms/strains | ||
|
| ||
| Mouse C57BL/6J | Charles River | Stock#027; RRID:IMSR_CRL:027 |
| Mouse BALB/c | Charles River | Stock#028; RRID:IMSR_CRL:028 |
| Mouse B6.SJL-Ptprca Pepcb/BoyJ | Jackson | Stock#002014; RRID:IMSR_JAX:002014 |
| Mouse CByJ.SJL(B6)-Ptprca/J | Jackson | Stock#006584; RRID:IMSR_JAX:006584 |
| Mouse B6.129S6-Del(3Cd1d2-Cd1d1)1Sbp/J | Jackson | Stock#008881; RRID:IMSR_JAX:008881 |
| Mouse C.129S2-Cd1tm1Gru/J | Jackson | Stock#003814; RRID:IMSR_JAX:003814 |
| Mouse B6.129S2-Cd28tm1Mak/J | Jackson | Stock#002666; RRID:IMSR_JAX:002666 |
| Mouse B6.129S4-Cd80tm1Shr Cd86tm2Shr/J | Jackson | Stock#003610; RRID:IMSR_JAX:003610 |
| Mouse B6.129P2(C)-Cd19tm1(cre)Cgn/J | Jackson | Stock#006785; RRID:IMSR_JAX:006785 |
| Mouse B6.Cg-Tg(Itgax-cre)1-1Reiz/J | Jackson | Stock#008068; RRID:IMSR_JAX:008068 |
| Mouse B6.Cg-Tg(Lck-cre)548Jxm/J | Jackson | Stock#003802; RRID:IMSR_JAX:003802 |
| Mouse B6 Foxn1-Cre-Tg | Georg Hollander | N/A |
| Mouse B6 CD28-tailless (TL) mutant | (Tai et al., 2005)23 | N/A |
| Mouse B6 Rag2-GFP-Tg | (Yu et al., 1999)45 | N/A |
| Mouse B6 CD28 mutant Knock-In (KI) | (Lio et al., 2010)24 | N/A |
| Mouse B6 B7flox | (Watanabe et al., 2017)36 | Stock#036705; RRID:IMSR_JAX036705 |
| Mouse BALB B7 DKO | (Williams et al., 2014)46 | N/A |
| Mouse BALB CD28 KO | (Williams et al., 2014)46 | N/A |
|
| ||
| Software and algorithms | ||
|
| ||
| FlowJo 10.8 | FlowJo, LLC | https://www.flowjo.com/ |
| R 4.1.1 | CRAN | https://cran.r-project.org/mirrors.html |
| RStudio 2021.09 | RStudio | https://rstudio.com/ |
| Seurat V4 | Hao et al., 202147 | https://satijalab.org/seurat/ |
| scRepetore 1.7.2 | Borcherding et al.,202048 | https://github.com/ncborcherding/scRepertoire/ |
| Graphpad Prism 8 | GraphPad Software | https://www.graphpad.com/scientific-software/prism/ |
|
| ||
| Other | ||
|
| ||
| Single Cell 5′ Library & Gel Bead Kit v1.1 | 10X Genomics | Cat# PN-1000165 |
EXPERIMENTAL MODEL AND SUBJECT DETAILS
C57BL/6 (B6) and BALB/c mice were purchased from Charles River. CB6F1 mice were bred in our facility. B6 CD45.1 congenic (B6.SJL-Ptprca Pepcb/BoyJ), BALB/c CD45.1 congenic (CByJ.SJL(B6)-Ptprca/J), B6 CD1d KO (B6.129S6-Del(3Cd1d2-Cd1d1) 1Sbp/J), BALB/c CD1d KO (C.129S2-Cd1tm1Gru/J), B6 CD28 KO (B6.129S2-Cd28tm1Mak/J), B6 B7.1/B7.2 double KO (DKO) (B6.129S4-Cd80tm1Shr Cd86tm2Shr/J), B6 CD19-Cre-KI (B6.129P2(C)-Cd19tm1(cre)Cgn/J), B6 CD11c-Cre-Tg (B6.Cg-Tg(It-gax-cre)1-1Reiz/J) and B6 Lck-Cre-Tg (B6.Cg-Tg(Lck-cre)548Jxm/J) mice were purchased from Jackson Laboratory. B6 Foxn1-Cre-Tg mice were generously provided by Dr. Georg Hollander. B6 CD28-tailless (TL) mutant mice23 were provided by Dr. Alfred Singer and Dr. Xuguang Tai. B6 Rag2-GFP-Tg mice45 and B6 CD28 mutant Knock-In (KI) mice24 were described previously. B6 B7flox mice were generated and described previously.36 BALB/c background CD28 KO and B7 DKO strains were generated at our facility by backcrossing B6 CD28 KO and B6 B7 DKO to BALB/c at least eight generations as previously described.46 Bone marrow chimera mice were generated by reconstitution with 107 total T cell–depleted BM cells from donor mice to irradiated (950 rad one time and 450 rad two times for B6 and BALB/c hosts, respectively) host mice i.v. Chimerism of radiation BM chimeras was analyzed 10 weeks after reconstitution with CD45.1 and CD45.2 donor cells and generally ranged between 20% and 80%. Chimeras that were more highly skewed to either CD45.1 or CD45.2 donor were excluded from analysis. 6- and 10-week-old female and male mice were used for all experiments. Mice were maintained in accordance with National Institutes of Health guidelines. All animal experiments were approved by the National Cancer Institute Animal Care and Use Committees.
V体育安卓版 - METHODS DETAILS
Flow cytometry
Cells were washed with FACS buffer (HBSS containing 0.2% BSA and 0.05% Azide), treated with anti-FcR (2.4G2), and then stained with the following Abs. Anti-CD4 (RM4-5)-BV510, anti-CD44 (IM7)-FITC, anti-TCRβ (H57-597)-PE/Cy7, anti-RORγt (Q31-378)-BV786, anti-T-bet (O4-46)-BV421, anti-PLZF (R17-809)-AF647, anti-CCR7 (4B12)-BV786, anti-IL-17A (TC11-18H10)-AF488, anti-IFNγ (XMG1.2)-FITC, anti-IL-4 (11B11)-BV421 were purchased from BD Biosciences. Anti-NK1.1 (PK136)-APC/eFluor780, anti-CD8α (53-6-7)-APC/eFluor780, anti-B7.1 (16-10A1)-APC, anti-B7.2 (GL1)-APC, anti-CD28 (37.51)-APC, anti-Egr2 (erongr2)-APC were purchased from eBioscience/Thermo Fisher. Anti-CD24 (M1/69)-AF594 and anti-CD69 (H1.2F3)-BV421 and anti-B220 (RA3-6B2)-AF700 were purchased from BioLegend. For detection of iNKT cells, PE-CD1d tetramer loaded with PBS57 (α-galactosylceramide analogue), obtained from the NIH Tetramer Core Facility, was used. For intracellular staining, Foxp3 staining kit (eBioscience/Thermo Fisher) was used for fixation and permeabilization according to manufacturer’s instructions and then stained with Abs for 30 min at 4°C. Data were collected with a FACS Fortessa (BD Biosciences) flow cytometer and analyzed with FlowJo (version 10.8) software (Tree Star).
In vitro cytokine assay
2 × 106 total thymocytes in 2 mL were placed in 24-well plates and treated with PMA (5 ng/mL) and Ionomycin (1000 ng/mL) for 4 h at 37°C. GolgiStop (0.67 μL/mL) and GolgiPlug (1 μL/mL) (eBioscience/Thermo Fisher) were added after 1 h of incubation. For intracellular staining, Foxp3 staining kit (eBioscience/Thermo Fisher) was used for fixation and permeabilization according to manufacturer’s instructions and cells then stained with Abs for 30 min at 4°C.
iNKT cell sort
BALB/c iNKT cells were magnetically enriched prior to flow cytometric sorting. Briefly, single cell thymocyte suspensions were incubated with PBS57-CD1d tetramer with 2.4G2 for 30 min at 4°C. Tetramer-bound cells were then enriched using EasySep PE-positive selection kit II (STEMCELL). The enriched cells were stained with FITC-CD44, TCRβ-APC, B220-AF700, CD24-AF594 Abs. Mature (B220−, TCRβ+ CD1d-tetramer + CD44high CD24low) and immature (B220− TCRβ+ CD1d-tetramer+, CD44low) iNKT cells were sorted separately with FACS Aria III (BD Bioscience). 25,000 cells of each sorted population were then mixed at a 50:50 ratio for scRNAseq in order to allow analysis of both mature and immature cells in the same sequencing runs.
Single-cell RNA-seq data processing and analysis
Approximately 10,500 cells were loaded in each capture lane of 10x Chromium to achieve a target of 6,000 captured cells, consistent with the 10X Genomics 5′ v2 Single Cell User Guide. Reverse transcription PCR was run overnight, and subsequent library preparation steps performed according to the User Guide. Three sequence runs have been performed, two NextSeq runs for GEX and one NextSeq run for GEX & TCR. Base calling was performed using RTA 3.9.2, demultiplexing was performed using Cell Ranger v6.1.1 (Bcl2fastq 2.20.0), and alignment was performed using Cell Ranger v6.1.1 (STAR 2.7.2a). Sequenced reads were aligned to the 10x Genomics provided mouse reference sequences (refdata-gex-mm10-2020-A for GEX and refdata-cell-ranger-vdj-GRCm38-alts-ensembl-5.0.0 for TCR). This Cell Ranger pipeline produced a gene expression matrix for each sample. Downstream analyses were performed using R version 4.1.1, RStudio version 2021.09.0 and the package Seurat version 4.0.5.47 Low-quality cells were filtered using the cutoffs nFeature_RNA >500 & nFeature_RNA <6,000 & percent.mt < 5. Data were normalized using NormalizedData function. IntegrateData function was used to integrate 6 data sets (WT n = 3, total 13,070 cells and B7 DKO n = 3, total 10,204 cells). For visualization and clustering, standard workflow was performed with RunPCA, RunUMAP, FindNeighbers and FindClusters funcitons. Cluster-enriched genes were identified by using the FindAllMarkers function with Wilcoxon-Rank sum test. Gene ontology (GO) analysis was performed by using DEenrichRPlot function. TCR Vβ gene usage of each NKT subset was quantified using scRepertoire.48
QUANTIFICATION AND STATISTICAL ANALYSIS
For statistical analysis, Prism GraphPad 8 software was used. Student’s t test with two-tailed distribution was performed for statistical analyses with a single comparison. For multiple comparisons, statistical analysis was performed with one-way ANOVA followed by Dunnett’s multiple comparison. p values <0.05 were considered statistically significant.
"V体育2025版" Supplementary Material
Highlights.
B7-CD28 co-stimulation is required for NKT1, NKT2, and NKT17 development
CD1d-expressing antigen-presenting T cells (APTCs) provide B7 co-stimulation
APTCs acquire B7 from other B7-expressing cells via CD28-dependent trogocytosis
Extracellular domain of CD28 on APTCs is sufficient to mediate B7 trogocytosis
V体育官网 - ACKNOWLEDGMENTS
We thank Alfred Singer, Yousuke Takahama, Xuguang Tai, and Karen Hathcock for their thoughtful comments and review of this manuscript. We thank Jonathan Green and Kelvin Lee for kindly providing CD28 mutant KI mice. We thank Xuguang Tai and Alfred Singer for providing CD28 mutant Tg mice. We thank Georg Hollander for providing Foxn1-Cre-Tg mice. We thank Michael Kelly and the National Cancer Institute Single Cell Analysis Facility for support of scRNAseq studies. Graphical abstract was created using BioRender (Bio.Render.com; publication and licensing rights agreement DE24JGT5KG). This work was supported by the Intramural Research Programs of the National Cancer Institute, National Institutes of Health.
"VSports" Footnotes
DECLARATION OF INTERESTS
The authors declare no competing interests.
SUPPLEMENTAL INFORMATION
Supplemental information can be found online at https://doi.org/10.1016/j.celrep.2022.111731.
REFERENCES
- 1.Bendelac A, Rivera MN, Park SH, and Roark JH (1997). Mouse CD1-specific NK1 T cells: development, specificity, and function. Annu. Rev. Immunol. 15, 535–562. 10.1146/annurev.immunol.15.1.535. [DOI] [PubMed] [Google Scholar]
- 2.Taniguchi M, Harada M, Kojo S, Nakayama T, and Wakao H (2003). The regulatory role of Valpha14 NKT cells in innate and acquired immune response. Annu. Rev. Immunol. 21, 483–513. 10.1146/annurev.immunol.21.120601.141057. [DOI] [PubMed] [Google Scholar]
- 3.Crosby CM, and Kronenberg M (2018). Tissue-specific functions of invariant natural killer T cells. Nat. Rev. Immunol. 18, 559–574. 10.1038/s41577-018-0034-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Coles MC, and Raulet DH (2000). NK1.1+ T cells in the liver arise in the thymus and are selected by interactions with class I molecules on CD4+CD8+ cells. J. Immunol. 164, 2412–2418. 10.4049/jimmunol.164.5.2412. [DOI] [PubMed] [Google Scholar]
- 5.Wei DG, Lee H, Park SH, Beaudoin L, Teyton L, Lehuen A, and Bendelac A (2005). Expansion and long-range differentiation of the NKT cell lineage in mice expressing CD1d exclusively on cortical thymocytes. J. Exp. Med. 202, 239–248. 10.1084/jem.20050413. [VSports在线直播 - DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zimmer MI, Colmone A, Felio K, Xu H, Ma A, and Wang CR (2006). A cell-type specific CD1d expression program modulates invariant NKT cell development and function. J. Immunol. 176, 1421–1430. 10.4049/jimmunol.176.3.1421. ["V体育官网" DOI] [PubMed] [Google Scholar]
- 7.Bai L, Constantinides MG, Thomas SY, Reboulet R, Meng F, Koentgen F, Teyton L, Savage PB, and Bendelac A (2012). Distinct APCs explain the cytokine bias of alpha-galactosylceramide variants in vivo. J. Immunol. 188, 3053–3061. 10.4049/jimmunol.1102414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Klein L, Kyewski B, Allen PM, and Hogquist KA (2014). Positive and negative selection of the T cell repertoire: what thymocytes see (and don’t see). Nat. Rev. Immunol. 14, 377–391. 10.1038/nri3667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chung B, Aoukaty A, Dutz J, Terhorst C, and Tan R (2005). Signaling lymphocytic activation molecule-associated protein controls NKT cell functions. J. Immunol. 174, 3153–3157. 10.4049/jimmunol.174.6.3153. [DOI (VSports手机版)] [PubMed] [Google Scholar]
- 10.Nichols KE, Hom J, Gong SY, Ganguly A, Ma CS, Cannons JL, Tangye SG, Schwartzberg PL, Koretzky GA, and Stein PL (2005). Regulation of NKT cell development by SAP, the protein defective in XLP. Nat. Med. 11, 340–345. 10.1038/nm1189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pasquier B, Yin L, Fondanèche MC, Relouzat F, Bloch-Queyrat C, Lambert N, Fischer A, de Saint-Basile G, and Latour S (2005). Defective NKT cell development in mice and humans lacking the adapter SAP, the X-linked lymphoproliferative syndrome gene product. J. Exp. Med. 201, 695–701. 10.1084/jem.20042432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kovalovsky D, Uche OU, Eladad S, Hobbs RM, Yi W, Alonzo E, Chua K, Eidson M, Kim HJ, Im JS, et al. (2008). The BTB-zinc finger transcriptional regulator PLZF controls the development of invariant natural killer T cell effector functions. Nat. Immunol. 9, 1055–1064. 10.1038/ni.1641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Savage AK, Constantinides MG, Han J, Picard D, Martin E, Li B, Lantz O, and Bendelac A (2008). The transcription factor PLZF directs the effector program of the NKT cell lineage. Immunity 29, 391–403. 10.1016/j.immuni.2008.07.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dose M, Sleckman BP, Han J, Bredemeyer AL, Bendelac A, and Gounari F (2009). Intrathymic proliferation wave essential for Valpha14+ natural killer T cell development depends on c-Myc. Proc. Natl. Acad. Sci. USA 106, 8641–8646. 10.1073/pnas.0812255106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mycko MP, Ferrero I, Wilson A, Jiang W, Bianchi T, Trumpp A, and MacDonald HR (2009). Selective requirement for c-Myc at an early stage of V(alpha)14i NKT cell development. J. Immunol. 182, 4641–4648. 10.4049/jimmunol.0803394. [DOI] [PubMed] [Google Scholar]
- 16.Lee YJ, Holzapfel KL, Zhu J, Jameson SC, and Hogquist KA (2013). Steady-state production of IL-4 modulates immunity in mouse strains and is determined by lineage diversity of iNKT cells. Nat. Immunol. 14, 1146–1154. 10.1038/ni.2731. [V体育ios版 - DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lee YJ, Wang H, Starrett GJ, Phuong V, Jameson SC, and Hogquist KA (2015). Tissue-specific distribution of iNKT cells impacts their cytokine response. Immunity 43, 566–578. 10.1016/j.immuni.2015.06.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wang H, and Hogquist KA (2018). How lipid-specific T cells become effectors: the differentiation of iNKT subsets. Front. Immunol. 9, 1450. 10.3389/fimmu.2018.01450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.White AJ, Lucas B, Jenkinson WE, and Anderson G (2018). Invariant NKT cells and control of the thymus medulla. J. Immunol. 200, 3333–3339. 10.4049/jimmunol.1800120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Greenwald RJ, Freeman GJ, and Sharpe AH (2005). The B7 family revisited. Annu. Rev. Immunol. 23, 515–548. 10.1146/annurev.immunol.23.021704.115611. [DOI] [PubMed] [Google Scholar]
- 21.Esensten JH, Helou YA, Chopra G, Weiss A, and Bluestone JA (2016). CD28 costimulation: from mechanism to therapy. Immunity 44, 973–988. 10.1016/j.immuni.2016.04.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Salomon B, Lenschow DJ, Rhee L, Ashourian N, Singh B, Sharpe A, and Bluestone JA (2000). B7/CD28 costimulation is essential for the homeostasis of the CD4+CD25+ immunoregulatory T cells that control autoimmune diabetes. Immunity 12, 431–440. 10.1016/s1074-7613(00)80195-8. [DOI] [PubMed] [Google Scholar]
- 23.Tai X, Cowan M, Feigenbaum L, and Singer A (2005). CD28 costimulation of developing thymocytes induces Foxp3 expression and regulatory T cell differentiation independently of interleukin 2. Nat. Immunol. 6, 152–162. 10.1038/ni1160. [DOI] [PubMed] [Google Scholar]
- 24.Lio CWJ, Dodson LF, Deppong CM, Hsieh CS, and Green JM (2010). CD28 facilitates the generation of Foxp3(−) cytokine responsive regulatory T cell precursors. J. Immunol. 184, 6007–6013. 10.4049/jimmunol.1000019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pobezinsky LA, Angelov GS, Tai X, Jeurling S, Van Laethem F, Feigenbaum L, Park JH, and Singer A (2012). Clonal deletion and the fate of autoreactive thymocytes that survive negative selection. Nat. Immunol. 13, 569–578. 10.1038/ni.2292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Watanabe M, Lu Y, Breen M, and Hodes RJ (2020). B7-CD28 co-stimulation modulates central tolerance via thymic clonal deletion and Treg generation through distinct mechanisms. Nat. Commun. 11, 6264. 10.1038/s41467-020-20070-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chung Y, Nurieva R, Esashi E, Wang YH, Zhou D, Gapin L, and Dong C (2008). A critical role of costimulation during intrathymic development of invariant NK T cells. J. Immunol. 180, 2276–2283. 10.4049/jimmunol.180.4.2276. [DOI] [PubMed] [Google Scholar]
- 28.Williams JA, Lumsden JM, Yu X, Feigenbaum L, Zhang J, Steinberg SM, and Hodes RJ (2008). Regulation of thymic NKT cell development by the B7-CD28 costimulatory pathway. J. Immunol. 181, 907–917. 10.4049/jimmunol.181.2.907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zheng X, Zhang H, Yin L, Wang CR, Liu Y, and Zheng P (2008). Modulation of NKT cell development by B7-CD28 interaction: an expanding horizon for costimulation. PLoS One 3, e2703. 10.1371/journal.pone.0002703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.McCaughtry TM, Wilken MS, and Hogquist KA (2007). Thymic emigration revisited. J. Exp. Med. 204, 2513–2520. 10.1084/jem.20070601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cowan JE, McCarthy NI, Parnell SM, White AJ, Bacon A, Serge A, Irla M, Lane PJL, Jenkinson EJ, Jenkinson WE, and Anderson G (2014). Differential requirement for CCR4 and CCR7 during the development of innate and adaptive alphabetaT cells in the adult thymus. J. Immunol. 193, 1204–1212. 10.4049/jimmunol.1400993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ueno T, Saito F, Gray DHD, Kuse S, Hieshima K, Nakano H, Kakiuchi T, Lipp M, Boyd RL, and Takahama Y (2004). CCR7 signals are essential for cortex-medulla migration of developing thymocytes. J. Exp. Med. 200, 493–505. 10.1084/jem.20040643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wang H, and Hogquist KA (2018). CCR7 defines a precursor for murine iNKT cells in thymus and periphery. Elife 7, e34793. 10.7554/eLife.34793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yu X, Fournier S, Allison JP, Sharpe AH, and Hodes RJ (2000). The role of B7 costimulation in CD4/CD8 T cell homeostasis. J. Immunol. 164, 3543–3553. 10.4049/jimmunol.164.7.3543. [DOI] [PubMed] [Google Scholar]
- 35.Benlagha K, Wei DG, Veiga J, Teyton L, and Bendelac A (2005). Characterization of the early stages of thymic NKT cell development. J. Exp. Med. 202, 485–492. 10.1084/jem.20050456. [DOI (V体育安卓版)] [PMC free article] [PubMed] [Google Scholar]
- 36.Watanabe M, Fujihara C, Radtke AJ, Chiang YJ, Bhatia S, Germain RN, and Hodes RJ (2017). Co-stimulatory function in primary germinal center responses: CD40 and B7 are required on distinct antigen-presenting cells. J. Exp. Med. 214, 2795–2810. 10.1084/jem.20161955. ["V体育官网" DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hwang I, Huang JF, Kishimoto H, Brunmark A, Peterson PA, Jackson MR, Surh CD, Cai Z, and Sprent J (2000). T cells can use either T cell receptor or CD28 receptors to absorb and internalize cell surface molecules derived from antigen-presenting cells. J. Exp. Med. 191, 1137–1148. 10.1084/jem.191.7.1137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sabzevari H, Kantor J, Jaigirdar A, Tagaya Y, Naramura M, Hodge J, Bernon J, and Schlom J (2001). Acquisition of CD80 (B7–1) by T cells. J. Immunol. 166, 2505–2513. 10.4049/jimmunol.166.4.2505. [DOI] [PubMed] [Google Scholar]
- 39.Lazarevic V, Zullo AJ, Schweitzer MN, Staton TL, Gallo EM, Crabtree GR, and Glimcher LH (2009). The gene encoding early growth response 2, a target of the transcription factor NFAT, is required for the development and maturation of natural killer T cells. Nat. Immunol. 10, 306–313. 10.1038/ni.1696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bettadapur A, Miller HW, and Ralston KS (2020). Biting off what can Be chewed: trogocytosis in Health, infection, and disease. Infect. Immun. 88, 009300–e1019. 10.1128/IAI.00930-19. ["VSports注册入口" DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sharrow SO, Mathieson BJ, and Singer A (1981). Cell surface appearance of unexpected host MHC determinants on thymocytes from radiation bone marrow chimeras. J. Immunol. 126, 1327–1335. [PubMed (V体育ios版)] [Google Scholar]
- 42.Sharpe AH, and Freeman GJ (2002). The B7-CD28 superfamily. Nat. Rev. Immunol. 2, 116–126. 10.1038/nri727. [DOI] [PubMed] [Google Scholar]
- 43.Ding L, and Shevach EM (1994). Activation of CD4+ T cells by delivery of the B7 costimulatory signal on bystander antigen-presenting cells (trans-costimulation). Eur. J. Immunol. 24, 859–866. 10.1002/eji.1830240413. [DOI] [PubMed] [Google Scholar]
- 44.Lee ST, Georgiev H, Breed ER, Ruscher R, and Hogquist KA (2021). MHC Class I on murine hematopoietic APC selects Type A IEL precursors in the thymus. Eur. J. Immunol. 51, 1080–1088. 10.1002/eji.202048996. ["V体育平台登录" DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yu W, Nagaoka H, Jankovic M, Misulovin Z, Suh H, Rolink A, Melchers F, Meffre E, and Nussenzweig MC (1999). Continued RAG expression in late stages of B cell development and no apparent re-induction after immunization. Nature 400, 682–687. 10.1038/23287. [DOI] [PubMed] [Google Scholar]
- 46.Williams JA, Zhang J, Jeon H, Nitta T, Ohigashi I, Klug D, Kruhlak MJ, Choudhury B, Sharrow SO, Granger L, et al. (2014). Thymic medullary epithelium and thymocyte self-tolerance require cooperation between CD28-CD80/86 and CD40-CD40L costimulatory pathways. J. Immunol. 192, 630–640. 10.4049/jimmunol.1302550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hao Y, Hao S, Andersen-Nissen E, Mauck WM 3rd, Zheng S, Butler A, Lee MJ, Wilk AJ, Darby C, Zager M, et al. (2021). Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587.e29. 10.1016/j.cell.2021.04.048. [VSports app下载 - DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Borcherding N, Bormann NL, and Kraus G (2020). scRepertoire: an R-based toolkit for single-cell immune receptor analysis. F1000Res. 9, 47. 10.12688/f1000research.22139.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
V体育官网入口 - Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Single-cell RNA-seq data have been deposited at the NCBI Gene Expression Omnibus (GEO: GSE198970) and are publicly available as of the date of publication. Accession numbers are listed in the key resources table. Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
This paper does not report original code.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
KEY RESOURCES TABLE.
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
|
| ||
| Antibodies | ||
|
| ||
| Anti-mouse CD4-BV510 (clone RM4-5) | BD Bioscience | Cat# 563106; RRID:AB_2687550 |
| Anti-mouse CD44-FITC (clone IM7) | BD Bioscience | Cat# 561859; RRID:AB_10894581 |
| Anti-mouse TCRβ-PE/Cy7 (clone H57-597) | BD Bioscience | Cat# 560729; RRID:AB_1937310 |
| Anti-mouse RORgt-BV786 (clone Q31-378) | BD Bioscience | Cat# 564723; RRID:AB_2738916 |
| Anti-mouse Tbet-BV421 (clone O4-46) | BD Bioscience | Cat# 563318; RRID:AB_2687543 |
| Anti-mouse PLZF-AF647 (clone R17-809) | BD Bioscience | Cat# 563490; RRID:AB_2738238 |
| Anti-mouse CCR7-BV786 (clone 4B12) | BD Bioscience | Cat# 564355; RRID:AB_2738765 |
| Anti-mouse IL-17A-AF488 (clone TC11-18H10) | BD Bioscience | Cat# 560220; RRID:AB_1645194 |
| Anti-mouse IFNγ-FITC (clone XMG1.2) | BD Bioscience | Cat# 554411; RRID:AB_395375 |
| Anti-mouse IL-4-BV421 (clone 11B11) | BD Bioscience | Cat# 562915; RRID:AB_2737889 |
| Anti-mouse NK1.1-APC/eF780 (clone PK136) | Thermo Fisher | Cat# 47-5941-82; RRID:AB_2735070 |
| Anti-mouse CD8α-APC/eF780 (clone 53-6-7) | Thermo Fisher | Cat# 47-0081-82; RRID:AB_469335 |
| Anti-mouse B7.1-APC (clone 16–10A1) | Thermo Fisher | Cat# 17-0801-82; RRID:AB_469417 |
| Anti-mouse B7.2-APC (clone GL1) | Thermo Fisher | Cat# 17-0862-82; RRID:AB_469419 |
| Anti-mouse CD28-APC (clone 37.51) | Thermo Fisher | Cat# 17-0281-82; RRID:AB_469374 |
| Anti-mouse Egr2-APC (clone erongr2) | Thermo Fisher | Cat# 17-6691-82; RRID:AB_11151502 |
| Anti-mouse B220-AF700 (clone RA3-6B2) | BioLegend | Cat# 103232; RRID:AB_493717 |
| Anti-mouse CD24-AF594 (clone M1/69) | BioLegend | Cat# 101834; RRID:AB_2565427 |
| Anti-mouse CD69-BV421 (clone H1.2F3) | BioLegend | Cat# 104545; RRID:AB_2686969 |
|
| ||
| Chemicals, peptides, and recombinant proteins | ||
|
| ||
| Mouse CD1d PBS-57 (PE-Labeled Tetramer) | NIH Tetramer core facility | Cat# 58281 |
| PMA | Sigma | Cat# P8139-1MG |
| Ionomycin | Sigma | Cat# 10634-1MG |
| EasySep PE-positive selection kit II | STEMCELL | Cat# 17684 |
| Foxp3/Transcription factor staining buffer set | Thermo Fisher | Cat# 00-5523-00 |
| GolgiPlug | BD Bioscience | Cat# 555029 |
| GolgiStop | BD Bioscience | Cat# 554724 |
|
| ||
| Deposited data | ||
|
| ||
| Single-cell RNA-seq data | This paper | GEO: GSE198970 |
|
| ||
| Experimental models: Organisms/strains | ||
|
| ||
| Mouse C57BL/6J | Charles River | Stock#027; RRID:IMSR_CRL:027 |
| Mouse BALB/c | Charles River | Stock#028; RRID:IMSR_CRL:028 |
| Mouse B6.SJL-Ptprca Pepcb/BoyJ | Jackson | Stock#002014; RRID:IMSR_JAX:002014 |
| Mouse CByJ.SJL(B6)-Ptprca/J | Jackson | Stock#006584; RRID:IMSR_JAX:006584 |
| Mouse B6.129S6-Del(3Cd1d2-Cd1d1)1Sbp/J | Jackson | Stock#008881; RRID:IMSR_JAX:008881 |
| Mouse C.129S2-Cd1tm1Gru/J | Jackson | Stock#003814; RRID:IMSR_JAX:003814 |
| Mouse B6.129S2-Cd28tm1Mak/J | Jackson | Stock#002666; RRID:IMSR_JAX:002666 |
| Mouse B6.129S4-Cd80tm1Shr Cd86tm2Shr/J | Jackson | Stock#003610; RRID:IMSR_JAX:003610 |
| Mouse B6.129P2(C)-Cd19tm1(cre)Cgn/J | Jackson | Stock#006785; RRID:IMSR_JAX:006785 |
| Mouse B6.Cg-Tg(Itgax-cre)1-1Reiz/J | Jackson | Stock#008068; RRID:IMSR_JAX:008068 |
| Mouse B6.Cg-Tg(Lck-cre)548Jxm/J | Jackson | Stock#003802; RRID:IMSR_JAX:003802 |
| Mouse B6 Foxn1-Cre-Tg | Georg Hollander | N/A |
| Mouse B6 CD28-tailless (TL) mutant | (Tai et al., 2005)23 | N/A |
| Mouse B6 Rag2-GFP-Tg | (Yu et al., 1999)45 | N/A |
| Mouse B6 CD28 mutant Knock-In (KI) | (Lio et al., 2010)24 | N/A |
| Mouse B6 B7flox | (Watanabe et al., 2017)36 | Stock#036705; RRID:IMSR_JAX036705 |
| Mouse BALB B7 DKO | (Williams et al., 2014)46 | N/A |
| Mouse BALB CD28 KO | (Williams et al., 2014)46 | N/A |
|
| ||
| Software and algorithms | ||
|
| ||
| FlowJo 10.8 | FlowJo, LLC | https://www.flowjo.com/ |
| R 4.1.1 | CRAN | https://cran.r-project.org/mirrors.html |
| RStudio 2021.09 | RStudio | https://rstudio.com/ |
| Seurat V4 | Hao et al., 202147 | https://satijalab.org/seurat/ |
| scRepetore 1.7.2 | Borcherding et al.,202048 | https://github.com/ncborcherding/scRepertoire/ |
| Graphpad Prism 8 | GraphPad Software | https://www.graphpad.com/scientific-software/prism/ |
|
| ||
| Other | ||
|
| ||
| Single Cell 5′ Library & Gel Bead Kit v1.1 | 10X Genomics | Cat# PN-1000165 |